
南湖新闻网讯(通讯员 胡健兵) 近日,国际植物学期刊Plant Biotechnology Journal在线发表了华中农业大学果蔬园艺作物种质创新与利用全国重点实验室/洪山实验室邓秀新院士团队柴利军教授组题为“Transposableelements cause the loss of self-incompatibility in citrus”的学术论文。研究系统证实控制自交不亲和S位点中S-RNase启动子附近MITE转座子的插入导致了芸香科金柑属(Fortunella)植物自交不亲和性状的丢失,且转座子插入事件在S-RNase介导的植物自交不亲和系统中具有广谱性,拓展了人们对植物自交不亲和向自交亲和性转变机制的新认识,建立了金柑属植物苗期分子标记辅助快速鉴定亲和性状技术体系,为柑橘种质创新与新品种培育提供了新思路,具有重要的理论及应用价值。
柑橘在世界范围内广泛栽培种植,其产业在国民经济及乡村振兴战略中具有重要意义。柑橘种质资源丰富,生殖模式多样,加之其童期较长、转化效率较低等问题,使得柑橘的生殖生物学研究,如自交不亲和机制解析相对滞后,极大限制了柑橘种质资源的开发和新种质培育。自交不亲和性(Self-incompatibility, SI)作为避免近亲繁殖和物种退化,保持物种遗传多样性重要的遗传机制,在被子植物中广泛存在。课题组前期研究表明,芸香科柑橘亚科属于以S-RNase介导的配子体型自交不亲和系统(Liang et al., 2020, Naure Plants),该系统在柑橘亚科不同属间的进化过程中经常丢失,即自交不亲和向自交亲和性转变。然而,对于这种向自交亲和转变的机制目前仍知之甚少。因此,搜集关键的柑橘自交亲和变异材料及发掘控制柑橘自交不亲和性转变的关键因子,并围绕其功能与作用机制进行解析,是进一步明确柑橘自交不亲和性作用机理及亲和性转变机制的关键,对柑橘新种质创制和新品种改良均具有重要的理论及实际意义。
SI系统遗传机制复杂,多由复等位的S位点所控制,S位点上包含两类在遗传上紧密连锁的即雌蕊S-RNase和雄蕊SLF基因。一旦这两类基因的连锁状态被打破或发生颠覆性突变,自交不亲和性就会被打破,最终会形成自交亲和物种。事实上,从强制性异交到自花受精的转变,在趋同进化中反复出现,很多自交亲和或自交物种均来自其自交不亲和祖先。而自交不亲和性的丢失是物种自交进化的关键,其对于自交不亲和系统背后复杂遗传机制的解析具有重要意义。
本研究以山金柑(Fortunella hindsiiSwingle)为研究对象,山金柑属于柑橘亚科金柑属植物,由于其具有极短的童期,播种当年可开花;超长的花期(武汉地区:5月初-9月底);易于遗传转化等特性(Zhuet al., Plant Biotechnology Journal, 2019, 17: 2199-2210),已经成为柑橘基因功能研究的模式材料。课题组前期经多年系统搜集和鉴定表明自然种群山金柑中包含自交不亲和及自交亲和两种生殖类型,为解析金柑属资源亲和转变机制奠定了材料基础。
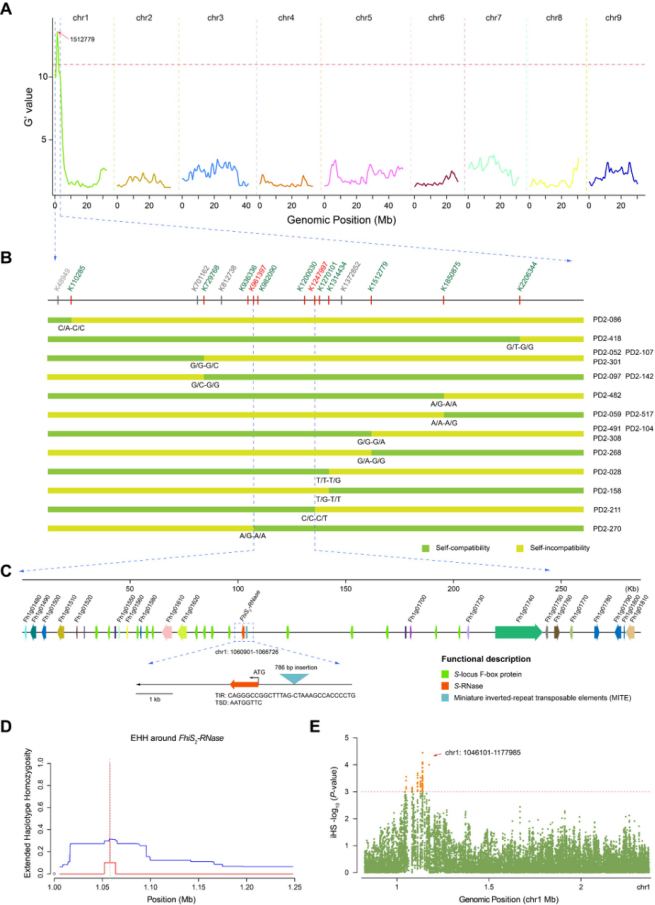
正向遗传学初定位S位点
为了探究山金柑(F. hindsii)由自交不亲和(Self-incompatibility, SI)向自交亲和(Self-compatibility,SC)转变的遗传基础,本研究基于已构建的PN02 × DB02杂交群体(其中PN02系为母本,为自交不亲和,基因型:S8S19;DB02系为父本,为自交亲和,基因型:S2S29),并对其杂交F1子代的SC表型进行了鉴定。经连续两年重复,共获得186个F1子代单株的自交亲和表型,其中SC与SI的分离比约为1:1(SC:SI = 98:88; χ2 = 0.54, P = 0.46)。基于全基因组测序分别选择32个SC和SI表型的F1单株用于BSA(Bulk Segregation Analysis, 混池分离群体分析)分析,在1号染色体亚端粒区发现一个与SC/SI相关的峰(0.83~2.38 Mb, G’ value = 11, P= 10-3)。基于KASP(Kompetitiveallele specific PCR, 竞争性等位基因特异性PCR)基因分型分析,从186株F1杂交子代中鉴定到18个在BSA定位区间发生重组的单株。依据其中的两个重组单株(PD02-211和PD02-270)及其自交亲和表型,明确了山金柑SC关键的遗传位点位于K961397和K1247997这两个标记之间,共286.6 kb的区间。该候选区间与控制柑橘自交不亲和性的S位点重叠,其中包含35个注释基因,包括14个F-box基因、1个S2-RNase基因(命名为FhiS2-RNase)和20个S位点侧翼区域的保守结构基因。
SC相关的位点在群体中通常处于强选择状态。针对这一候选区域,本研究计算了山金柑自交亲和自然群体的EHH(Extended Haplotype Homozygosity, 扩展单倍型纯和度),测量了携带特定核心等位基因单倍型的衰减作为距离的函数,并获得了每个SNP的iHS(integratedhaplotype score, 综合单倍型得分)统计数据,以寻找近期有正选择迹象的基因组区域。在FhiS2-RNase的基因组区域发现了一个强烈的正选择信号,该区域包含75个显著的SNPs(P = 10-3),并与上述35个注释基因中的13个相关。此外,杂交群体F1子代SC表型与S2-RNase基因型密切相关。序列比对结果表明,S2-RNase基因编码区在不同柑橘属间和种间存在几个同义和非同义突变,其中FhiS2-RNase编码的氨基酸序列相对于其他5个不同属间和种间的S2-RNase基因(PtrS2-RNase, CmeS2-RNase, AbuS2-RNase,CgrS2-RNase和CreS2-RNase)相比,在C端多出12个氨基酸残基,但这并未改变其保守结构域(C1-C5)。
负频率依赖选择是SI的一个关键特征,它决定了S位点等位基因的进化动态。本研究对来自中国5个省的46份山金柑核心种质资源的基因型和自交不亲和表型进行了鉴定。其中S2-RNase基因型的占比最高(30/46),高达33.7%。值得注意的是,所有包含S2-RNase基因型的种质均为自交亲和。为了检测山金柑SC和SI种群之间的正选择特征,本研究对其进行了XP-EHH(Cross-Population Extended Haplotype Homozygosit, 跨群体扩展单倍型纯和度)分析。在3条染色体上检测到4个选择信号,共获得34个假定有利的正选择基因,其中17个基因与KASP基因分型结果重叠,表明包括FhiS2-RNase在内的这些基因在自交亲和山金柑品系中被强烈选择。
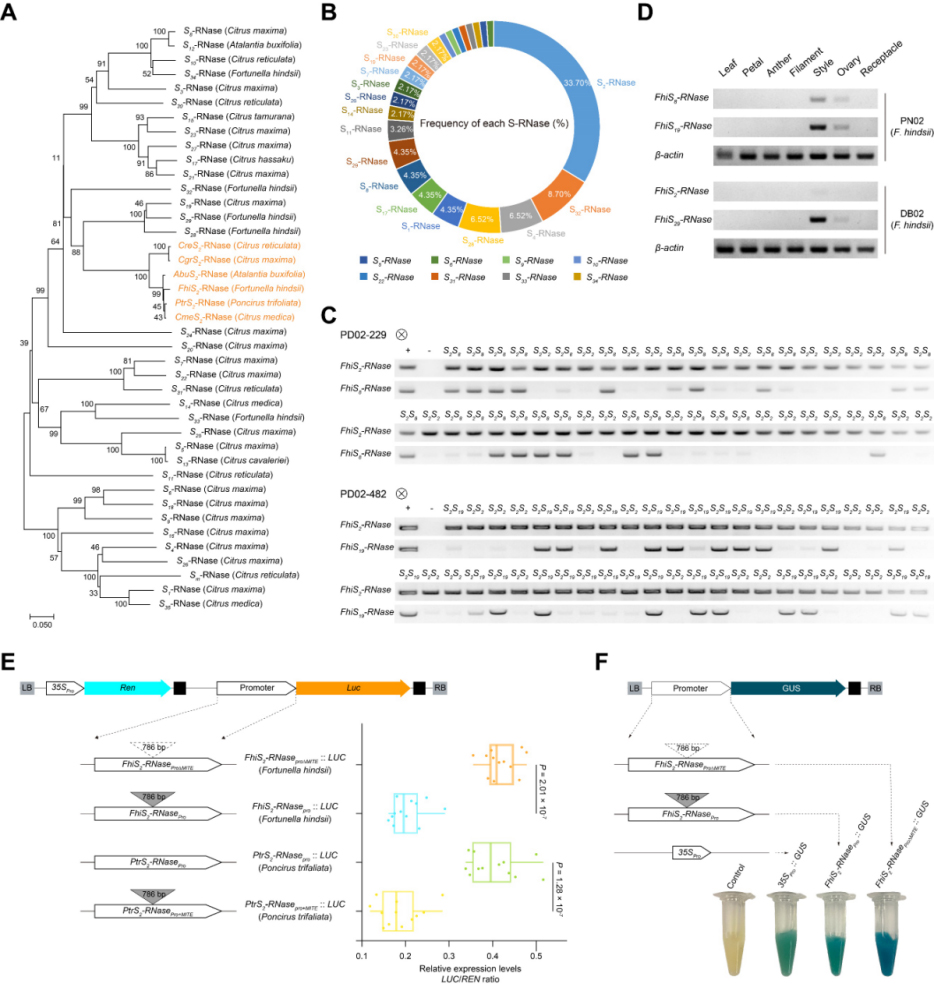
MITE转座子对S-RNase基因表达影响
为了进一步评估FhiS2-RNase是否是山金柑从SI向SC转变的候选基因,基于卡方检验(χ2-square)分析了自交亲和山金柑自交子代(S1)中S基因型的分离比率。自花授粉PD02-229(S2S8)的子代S单倍型,并不是按照1:2:1的比例分离,而是按照1:1(S2S2:S2S8 = 62:63)的比例分离,且在S1子代中并未出现S8S8的基因型,这一结果与S2位点的异常相一致(1:1, χ2= 0.01, P = 0.93)。此外,来自于自花授粉PD02-482(S2S19)的116个子代单株的基因型分离比也遵从1:1(S2S2:S2S19 = 55:61; 1:1, χ2 =0.31, P = 0.58),并且其S1子代中也未出现S19S19基因型。这些数据表明,只有携带S2等位基因的花粉,而不是携带S8或S19等位基因的花粉,能够传递给S1子代,结果进一步支持了FhiS2-RNase基因型与山金柑SI的丢失密切相关。
实时半定量PCR(qRT-PCR)结果显示,自交亲和山金柑DB02系花柱组织中FhiS2-RNase基因的表达显著被抑制,表明FhiS2-RNase的这些改变可能是导致山金柑自交亲和的原因。在对山金柑和枳(ZK8,Poncirus trifoliata)的S2位点序列进行比对时,发现位于FhiS2-RNase基因启动子上游1071 bp处存在一个786 bp的插入序列。由于不同柑橘属的S2-RNase基因非编码区存在显著差异,只有枳(ZK8)和山金柑在该区域具有较高的同源性。基于P-MITE数据库和RepeatMasker软件预测结果,确定了这786 bp的插入序列是一个属于hAT(hobo, Ac and Tam3)家族的MITE(Miniature inverted-repeat transposable elements)转座子。
为了确定该MITE转座子的插入是否影响了FhiS2-RNase基因的表达,将来自山金柑的不同启动子片段插入到pGreenⅡ 0800-LUC和pKGWFS7载体中,构建重组质粒。随后,将这些重组载体转入到山金柑DB02系的愈伤组织中,并检测了LUC(luciferase,荧光素酶报告基因)的表达。去掉了MITE转座子后的重组载体(FhiS2-RNaseproΔMITE::LUC),其启动子活性得到了显著增强(P = 3.36 × 10-5),这一结果与GUS组织化学染色结果一致。此外,当在枳S2-RNase基因启动子相同位置上插入该MITE转座子(PtrS2-RNasepro+MITE::LUC),其PtrS2-RNase启动子活性则显著降低(P = 9.59 × 10-7)。这些数据显示,MITE转座子的插入可以调控山金柑中FhiS2-RNase基因的表达水平,同时786 bp的MITE转座子插入可能是山金柑SI丢失的原因。
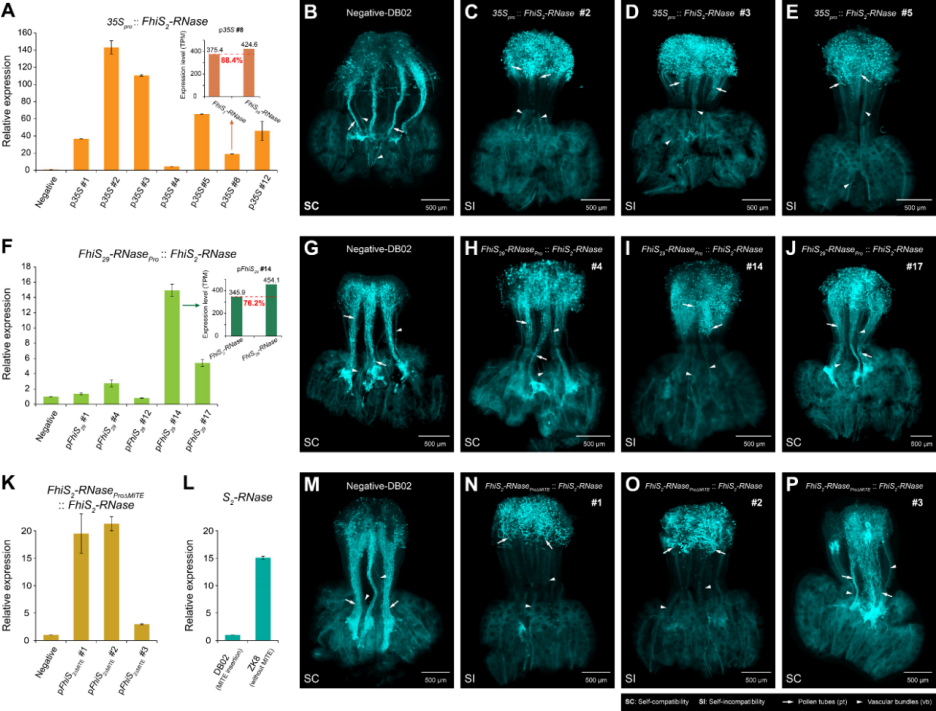
MITE转座子活体功能验证
多年生木本植物开展活体基因功能验证耗时长,难度大,但为了确认FhiS2-RNase启动子中的MITE转座子插入是否导致其低水平表达,本研究构建了去除786 bp MITE插入的互补转基因株系(FhiS2-RNaseproΔMITE::FhiS2-RNase-DB02)。在获得的3个稳定开花的转基因株系中,2个株系FhiS2-RNase基因的相对表达量提高了约20倍,苯胺蓝染色结果显示,其花柱可以排斥自我花粉管的生长,表明与对照相比,SI表型已经得到恢复。此外,本研究还发现S2-RNase (PtrS2-RNase)在ZK8 (P. trifoliata, S2S31)花柱组织中的表达量显著高于DB02 (F. hindsii, FhiS2-RNase),其启动子与FhiS2-RNase相比缺少了786 bp的MITE转座子插入。这些结果表明,786 bp的MITE转座子插入抑制了FhiS2-RNase的表达,并导致了山金柑SI的丢失。
进一步对柑橘家族S位点分析显示,MITE转座子插入事件在柑橘不同属间和种间S-RNase等位基因附近频繁发生,而且特定的MITE转座子插入会影响宿主基因的表达(FhiS2-RNase,PtrS31-RNase及CreS31-RNase),在柑橘SI丢失中起重要作用。此外,在其他常见配子体自交不亲和(gametophyticself-incompatibility)物种中也观察到MITE转座子在S-RNases等位基因附近插入,以上结果表明,基于S-RNases的SI系统中,MITE介导的SI丢失普遍存在。
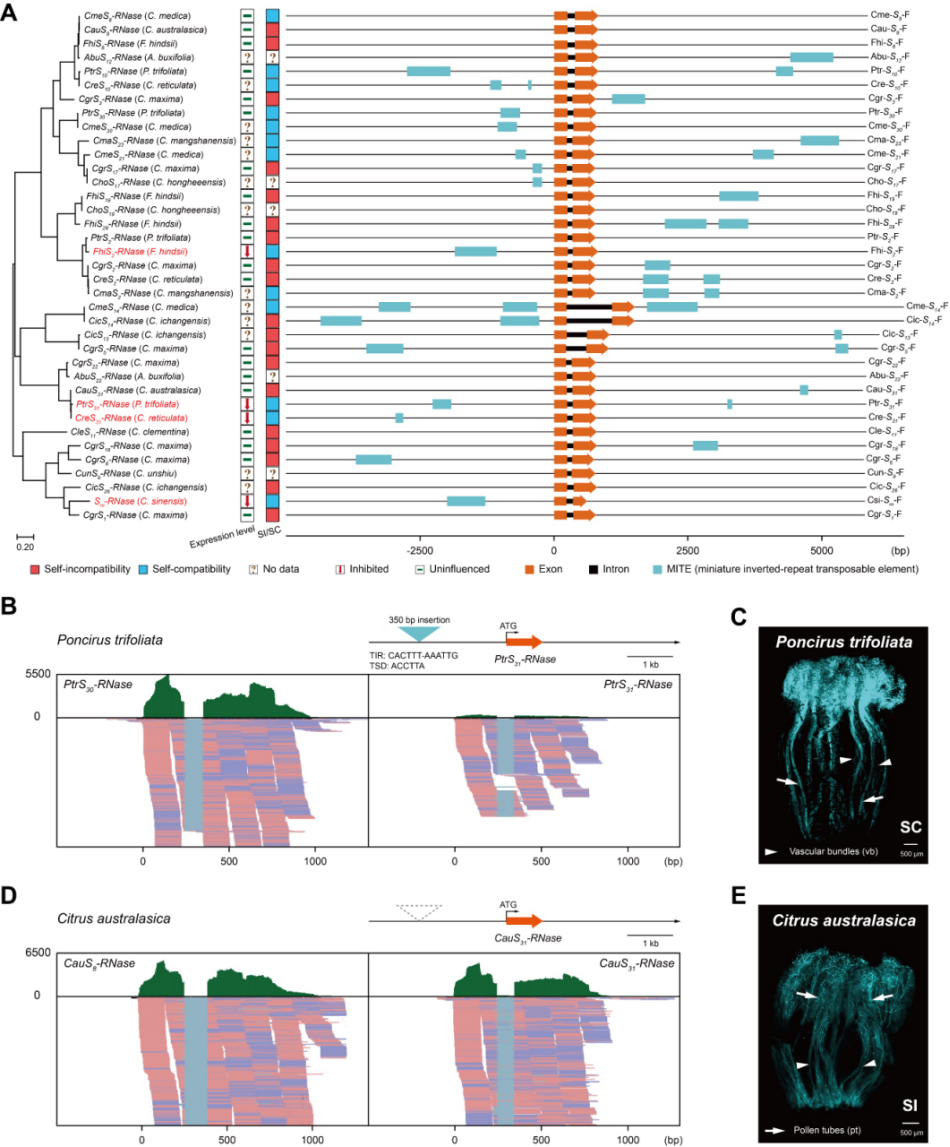
MITE转座子广谱性存在
SI的丢失在适应性进化过程中起着至关重要的作用,随着越来越多的功能性转座子(TEs,Transposable elements)被发现,转座子对物种分化和驯化的影响将会进一步得到阐释。本研究对S-RNases相关的MITE转座子插入在自交进化中的功能分析拓宽了人们对自交不亲和系统遗传基础的理解,也为后续S-RNase和SLF特异性识别的生化基础及共同进化研究奠定了坚实的基础。
华中农业大学博士后胡健兵为论文第一作者,果蔬园艺作物种质创新与利用全国重点实验室、洪山实验室固定研究人员柴利军教授为论文通讯作者。华中农业大学博士研究生刘晨晨、杜泽臻及已毕业硕士生郭福蓉、宋丹及韦壮敏(现工作单位广西壮族自治区亚热带作物研究所);已毕业博士王楠(现单位中国农业科学院深圳农业基因组研究所);湖南省农业科学院园艺所陈鹏;华中农业大学果蔬园艺作物种质创新与利用全国重点实验室公共平台石春梅工程师;华中农业大学林宗成教授、徐强教授、邓秀新院士及英国伯明翰大学Vernonica E. Franklin-Tong教授、英国阿伯里斯特维斯大学Maurice Bosch教授参与了本研究工作。本研究得到国家重点研发计划,国家自然科学基金及国家现代农业产业技术体系等项目资助。
链接:https://onlinelibrary.wiley.com/doi/full/10.1111/pbi.14250
审核人 柴利军